细胞质基质的功能
- 蛋白质的修饰
- 控制蛋白质的寿命
- 降解变性和错误折叠的蛋白质
- 帮助变性或错误折叠的蛋白质重新折叠,形成正确的分子构象
- 这一功能靠热休克蛋白(heat shock protein,hsp)完成,其作为分子伴侣(molecular chaperone)发挥作用,协助蛋白质合成、分选、折叠、装配。
- 内质网中的结合蛋白(binging protein,Bip),属于Hsp70家族的分子伴侣
内质网功能
脂质、蛋白质、糖类合成
- 蛋白质合成
- 蛋白质都在核糖体合成,起始于游离核糖体
- 主要合成分泌蛋白(抗体)、膜整合蛋白、内膜系统的细胞器内的可溶性驻留蛋白(溶酶体中的的酸性水解酶)
- 脂质的合成:所有的磷脂和胆固醇(sER)
- 磷脂转运的方式:
- 转运到高尔基体、溶酶体和细胞膜以出芽的方式
- 转运到线粒体、过氧化物酶体磷脂转换蛋白
- 磷脂转运的方式:
- 蛋白质的修饰与加工:糖基化(N-连接的糖基化在内质网)
- 新生多肽的折叠与组装:蛋白二硫键异构酶、结合蛋白(binding protein,Bip)
- 储存Ca2+离子
- 确定膜蛋白的方向性
高尔基体的功能
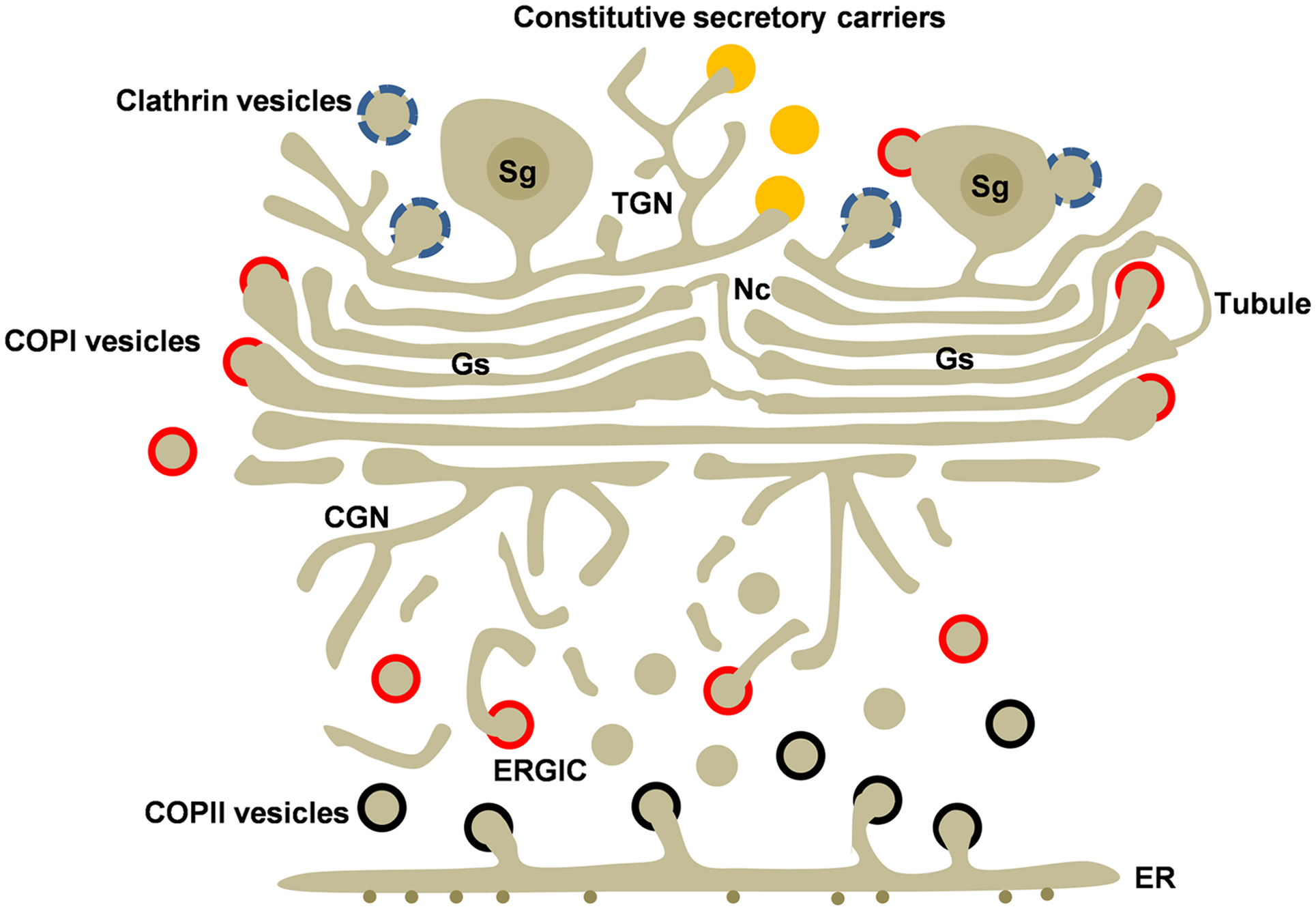
图片alt
内质网合成蛋白进行加工、分类、包装、装运;合成糖类
- 内质网合成的蛋白质和脂类进入CGN区域,带有KDEL和HDEL信号的蛋白返回内质网
- TGN区域、参与蛋白质的转运、包装
- 发生O-连接的糖基化修饰
- 蛋白质自身所携带的分选信号决定了蛋白质的去向,如:溶酶体蛋白的M6P分选途径
- 分泌性蛋白的分泌过程:rER合成蛋白质、进入ER腔、COPⅡ运输泡、进入CGN区、在中间膜囊加工、在TGN区形成小泡、小泡与质膜融合排出
- 硫酸根转移到肽链中的酪氨酸残基上
蛋白质糖基化修饰
- N-连接糖基化和O-连接糖基化比较
| 特征 | N-连接 | O-连接 |
|---|---|---|
| 合成部位 | 粗面内质网 | 主要在高尔基体 |
| 合成方式 | 来自同一个寡糖前体 | 一个个单糖加上去 |
| 与之结合的氨基酸残基 | 天冬氨酸 | 丝氨酸、苏氨酸、羟脯氨酸、羟赖氨酸 |
| 最终长度 | 至少5个糖基 | 1~4个糖基 |
| 第一个糖残基 | N-乙酰基葡萄糖胺 | N-乙酰半乳糖胺 |
溶酶体
-
阶段:
- 初级溶酶体
- 次级溶酶体
- 残余小体
-
功能:
- 清除作用
- 防御功能
- 营养作用
- 清除赘生组织或退行性变化细胞
- 参与分泌调节
- 精子顶体
-
溶酶体的发生:依赖于M6P的溶酶体酶分选途径
- 在rER上的核糖体合成溶酶体酶蛋白
- 进入内质网腔,进行N-连接的糖基化修饰
- 在高尔基体,顺面膜囊,寡糖链上的甘露糖残基磷酸法形成M6P(甘露糖-6-磷酸)
- 与高尔基体反面膜囊和反面网状结构上的M6P受体结合
- 以出芽的方式转运至运输小泡
- 运输小泡与溶酶体前体融合,M6P受体返回高尔基体
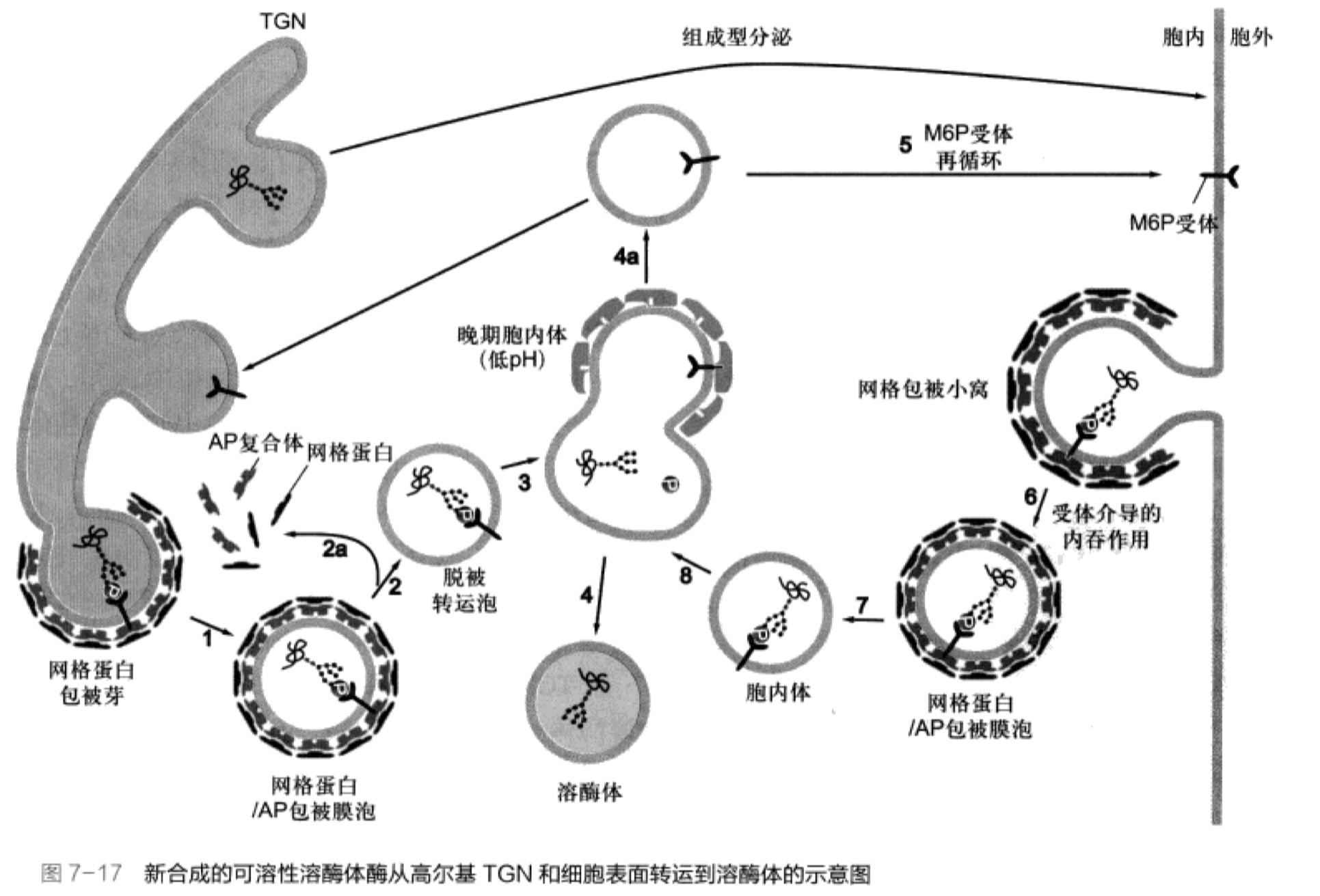
图片alt
-
溶酶体与过氧化物酶体比较
| 特征 | 溶酶体 | 过氧化物酶体 |
|---|---|---|
| 形态 | 球形、无晶格状结构 | 球形,有晶格状结构 |
| 酶种类 | 酸性水解酶 | 氧化酶 |
| pH | 5左右 | 7作用 |
| 是否需要O2 | 不需要 | 需要 |
| 功能 | 细胞内消化作用 | 多种功能 |
| 发生 | 内置网合成,高尔基体出芽形成 | 细胞质基质合成,分裂组装形成 |
| 识别的标志酶 | 酸性水解酶 | 过氧化氢酶 |
电镜下可用于识别过氧化物酶体的主要特征是:由尿酸氧化酶形成的晶格状结构
溶酶体、过氧化物酶体(微体)植物细胞称为乙醛酸循环体都是异质性细胞器。所谓异质性是指,溶酶体的形态大小,甚至其所包含的水解酶种类可能不同
蛋白质的分选
- 指导分泌蛋白在糙面内质网合成的决定因素:
- N端的信号肽(signal peptide)
- 信号识别颗粒(signal recognition particle,SRP)
- 内质网膜上信号肽识别颗粒受体(docking protein,DP)
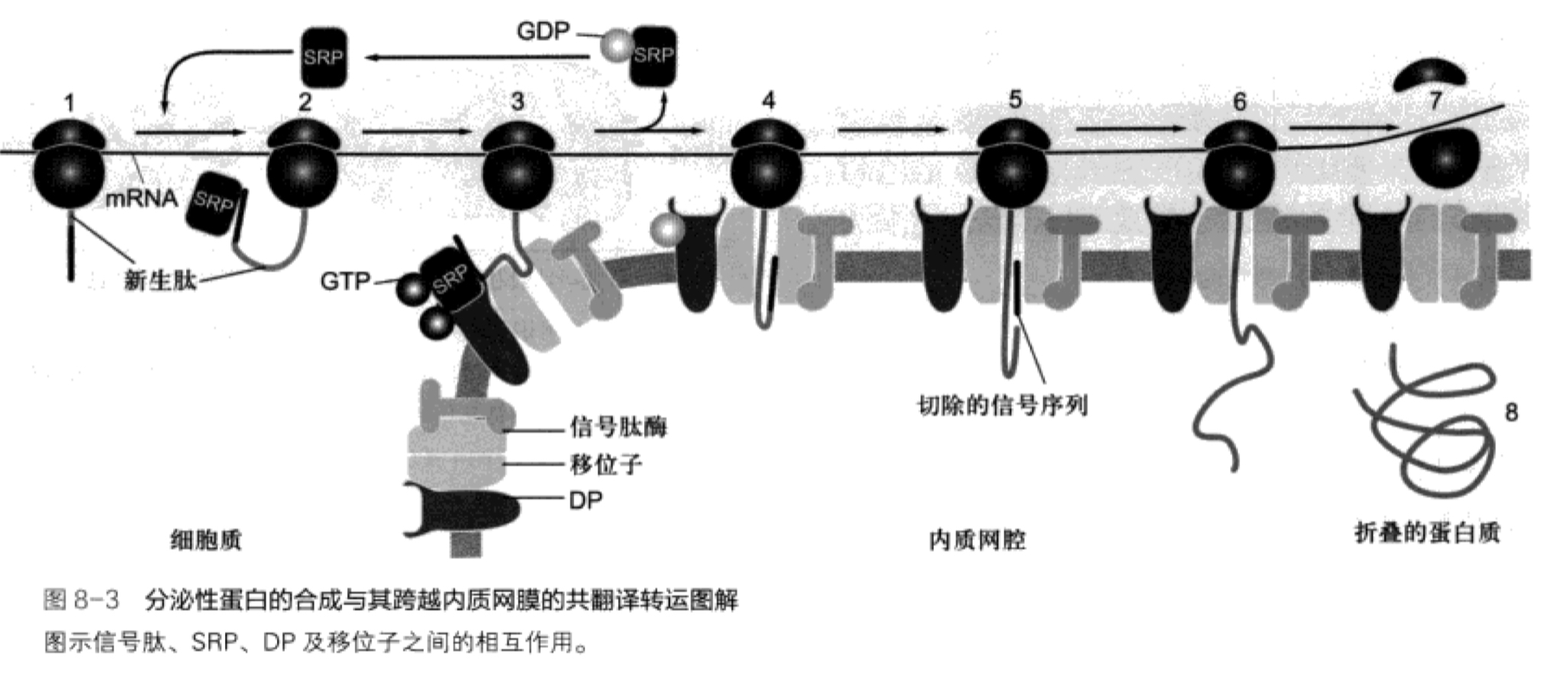
图片alt
- 后翻译转运途径
- 游离核糖体合成
- 共翻译转运途径
- 在游离核糖体起始,由信号肽和SRP(信号肽识别颗粒)引导转移到糙面内质网
膜泡运输
- COPⅡ 包被小泡
- 高尔基体➡️内质网
- COPⅠ包被小泡
- 内质网➡️高尔基体
- 网格蛋白/接头蛋白
- 高尔基体TGN➡️脂膜、包内体、溶酶体